
Introduction
The kidney bean, or Phaseolus vulgaris L., is a significant grain legume prized for its high protein content, dietary fibre and micronutrients, and it makes a significant contribution to food and nutrition security in many regions.1-4 In developing countries, where plant‑based foods are a major protein source, common bean plays a particularly important role in human diets.3-5 However, its genetic improvement is constrained by relatively narrow genetic diversity and autogamous reproduction,6,7 which limit the creation and exploitation of new recombinants through conventional breeding.
Mutation breeding provides a useful supplementary strategy for expanding genetic variability in such crops and has been widely applied in grain legumes.8,9 Physical mutagens such as gamma rays and chemical mutagens such as Sodium azide and EMS are frequently used to induce point mutations and chromosomal alterations that can generate a variety of phenotypic variants.8-10 In legumes, mutagenic treatments have been demonstrated to impact germination, vegetative growth and reproductive performance, and to induce chlorophyll‑deficient and morphological mutants that serve as useful genetic markers.10-14
Despite this progress, comparative studies that simultaneously evaluate physical and chemical mutagens across different kidney bean genotypes under Indian agro‑climatic conditions remain limited.6,9 Moreover, genotype‑specific responses to different mutagenic agents are not fully characterized,8,10 especially in cultivars that have been acclimated to the area and are crucial to regional cropping systems.
Thus, the purpose of this study was to compare the impact of gamma radiation, EMS and sodium azide on germination, early growth, mutation frequency and yield‑related traits in two kidney bean cultivars, Varun and Waghya, adapted to Indian conditions. The specific objective was to identify mutagenic treatments capable of inducing useful phenotypic variability, while documenting cultivar‑specific differences in mutability and tolerance that can guide future mutation breeding programmes.
Materials and Methods
Plant material
Seeds of kidney bean (Phaseolus vulgaris L.) cultivars Varun (ACPR-94040) and Waghya (HPR-35) were used in the experiment. Healthy, uniform and well-dried seeds with approximately 10% moisture content was chosen selected for mutagenic treatment.
Mutagenic treatments
Gamma irradiation
Seeds were exposed to gamma rays at doses of 10kR, 15kR, 20kR and 25 kR using a cobalt‑60 source with a dose rate of 24,578 rads h⁻¹. The radiation was administered at the Government Institute of Science’s Department of Biophysics in Aurangabad, Maharashtra, India.
Chemical mutagen treatments
The seeds underwent surface sterilization using 0.1% mercuric chloride for 1 min and then thoroughly cleaned with distilled water. Distilled water was used to pre-soak them for 6 h before chemical mutagen treatment. EMS was applied at concentrations of 0.10, 0.15, 0.20 and 0.25% for 4 h, and sodium azide was applied at 0.010, 0.015, 0.020 and 0.025% for 6 h. Every chemical treatment was carried out with constant shaking at 25 ± 2°C, and the mutagen solution volume was kept up at three times the seed volume. After treatment, The seeds were cleaned under running water and post-soaked in distilled water for 2 h before drying.
Experimental design and field layout
Each treatment consisted of 200 seeds. For germination testing, 25 seeds were placed on moistened blotting paper in Petri dishes, and another 25 seeds were grown in pots for seedling measurements. The remaining 150 seeds were planted in the field in three replications using a randomized block design, each containing 50 seeds. The field study was carried out in the kharif season at the Department of Botany’s experimental farm of Arts, Commerce and Science College, Sonai, Tal. Newasa, Dist. Ahilyanagar, India. Seeds were sown at 20 cm intra‑row and 45 cm inter‑row spacing, and Standard agronomic procedures were adhered to the season.
Observations recorded
The M₁ generation was assessed for germination percentage, seedling height, leaf morphological mutations, chlorophyll chimeras, plant survival, pod length, number of pods/plants, seeds/pod, seed size & seed coat colour.
Germination was recorded when both plumule and radicle emerged. Ten randomly chosen seedlings from each treatment were used to measure the height of the seedlings on the tenth day following germination. Leaf morphological mutations were documented as plants showing altered leaf shape, size or margin, while chlorophyll chimeras were recorded as plants showing variegated or chlorophyll‑deficient sectors. Plant survival was recorded at physiological maturity and stated as a proportion of total plants established in the field.
Mutation frequency for leaf morphological mutations and chlorophyll chimeras was calculated as:
Mutation frequency (%) = (Number of mutant plants / Total plants observed) × 100
For each treatment, all surviving M₁ plants were examined, and only clearly distinguishable mutants were comprised in the counts.
Pod length was measured at harvest. Pods / plant and seeds/pod were counted at maturity to measure the influence of mutagenic treatments on reproductive performance. Seed size was visually classified as medium, small–medium or small in Varun, and medium–large, medium, medium–small or small in Waghya. Seed coat colour variants were recorded qualitatively and compared with the control phenotype.
Statistical analysis
Data on germination percentage, seedling height, leaf morphological mutations, chlorophyll chimeras, plant survival, pod length, pods/plant and seeds/pod were summarized using descriptive statistics. The results are presented as mean ± standard error (SE) based on three replications under a randomized block design (RBD).
Safety considerations
All procedures involving mercuric chloride and sodium azide were performed in a fume hood while using gloves, laboratory coats and eye protection. Residual mutagen solutions were collected separately and disposed of according to institutional hazardous‑waste protocols to minimize environmental risk.
Results
Germination and seedling growth
All mutagenic treatments reduced germination and seedling height in both cultivars compared with the control, and the reductions generally intensified with increasing dose (Figure 1.and 2.). In Varun, germination declined from 94.10% in the control to 60.25% under SA 0.020%, while seedling height decreased from 12.48 cm to 5.15 cm at the same dose. In Waghya, germination declined from 84.10% to 60.35% and seedling height from 12.08 cm to 5.08 cm under SA 0.020%. Sodium azide thus caused the strongest inhibition, followed by EMS, whereas gamma rays produced comparatively milder reductions in both traits. These dose‑dependent responses indicate clear phytotoxic effects of the mutagenic treatments on early seedling performance, consistent with observations in other legume crops.10,12,13,15
Leaf morphological mutations
Leaf morphological mutations increased with increasing dose in both cultivars (Figure 3.). In Varun, the maximum frequency was detected under SA 0.020% (17.95%), whereas in Waghya it was highest under EMS 0.20% (17.05%). The variants included plants with altered leaf size, changes in lamina shape and modifications in margin pattern. These morphological anomalies are consistent with previous studies of increased leaf mutation frequency under EMS and SA treatments in legumes and French beans, and they show significant mutagenic harm in the M₁ generation.10-13
Chlorophyll chimeras
Chlorophyll‑deficient chimeras were detected in both cultivars and their frequency increased with mutagen dose (Figure 4.). SA treatments caused the highest frequencies of chimeras, followed by EMS and gamma rays, with maximum values recorded at SA 0.020% in both cultivars. The chimeras included albina, chlorina and xantha‑type sectors in seedlings. As these chimeras are typically somatic and unstable in M₁, they serve as sensitive indicators of mutagenic activity rather than stable genetic changes, in accordance with the well‑established role of chlorophyll alterations as biomarkers for mutagenic effectiveness.14
Plant survival
Plant survival declined with increasing dosage in both cultivars. Varun showed a decline from 92.30% in the control to 73.10% under SA 0.020%, while Waghya declined from 85.45% to 70.10% at the same dose. Sodium azide again produced the strongest biological impact, while gamma rays caused moderate reductions in survival. Despite these declines, survival remained relatively high at lower mutagen doses, indicating that some treatments were severe but not lethal and may still allow recovery of useful mutants. Similar reductions in survival under mutagenic stress have been reported in other bean and legume studies.8,9,15
Yield‑related traits
Pod length, pods/plant and seeds/pod were reduced under mutagenic treatments in both cultivars. In Varun, pod length decreased from 15.4 cm in the control to 11.5 cm under EMS 0.25%, while pods/plant declined from 9.5 to 6.8 & seeds/pod from 5.6 to 4.3. In Waghya, pod length decreased from 16.4 cm to 12.7 cm under SA 0.020%, pods/plant declined from 9.3 to 6.9 and seeds/pod from 5.3 to 4.5. Waghya consistently maintained slightly better reproductive performance than Varun across most treatments, indicating comparatively greater tolerance to mutagenic stress. The greater sensitivity of pod length and pod number compared with seeds/pod advises that early reproductive development is more vulnerable to mutagen introduction, which is consistent with earlier reports on mutagen effects on reproductive traits in pulses and beans.16-18
Seed size and seed coat colour
Seed size gradually shifted from larger to smaller categories with increasing mutagen dose. In Varun, seed size varied from medium in the control and low‑dose treatments to small or small–medium at higher EMS and SA concentrations. In Waghya, seed size changed from medium–large or medium in the control and lower doses to medium–small or small under stronger mutagenic treatments. Seed coat colour also varied among treatments, including cream, pale brown, light yellow‑brown and light red‑brown shades, but these changes were irregular and not clearly dose‑dependent. As expected for the M₁ generation, such seed coat colour variants are likely to represent somatic or physiological changes rather than stable, heritable mutations, and true seed coat colour mutants are more reliably identified in M₂ and later generations.9,19
Table 1: Effect of EMS, sodium azide and gamma irradiation on M₁ germination, growth, mutation frequencies and yield-related traits in kidney bean variety ‘Varun’
| Treatment | Dose | Seed germination (%) |
Seedling height (cm) |
Leaf morph. changes (%) |
Chlorophyll chimeras (%) |
Plant survival (%) | Pod length (cm) | Pods/ plant |
Seeds/ pod |
| Control | — | 94.10 ± 0.42 |
12.48 ± 0.66 |
— | — | 92.30 ± 0.51 |
15.4 ± 0.38 |
9.5 ± 0.25 |
5.6 ± 0.12 |
| EMS | 0.10% | 84.20 ± 0.61 |
8.82 ± 0.39 |
9.90 ± 0.28 |
4.45 ± 0.14 |
86.52 ± 0.41 |
14.1 ± 0.40 |
8.2 ± 0.22 |
5.1 ± 0.10 |
| EMS | 0.15% | 80.45 ± 0.74 |
7.81 ± 0.44 |
12.10 ± 0.54 |
5.62 ± 0.58 |
80.45 ± 0.48 |
13.2 ± 0.37 | 7.6 ± 0.20 |
4.8 ± 0.11 |
| EMS | 0.20% | 77.10 ± 1.08 |
6.05 ± 0.41 |
14.02 ± 0.47 |
6.58 ± 0.36 |
75.80 ± 0.68 |
12.0 ± 0.33 |
7.0 ± 0.19 |
4.4 ± 0.09 |
| EMS | 0.25% | 76.85 ± 1.12 |
5.92 ± 0.39 |
14.35 ± 0.51 |
6.75 ± 0.42 |
75.10 ± 0.71 |
11.5 ± 0.30 |
6.8 ± 0.18 |
4.3 ± 0.09 |
| SA | 0.010% | 70.85 ± 0.82 |
7.05 ± 0.11 |
12.75± 0.66 | 7.82 ± 0.30 |
81.70 ± 0.77 |
13.3 ± 0.35 |
7.9 ± 0.21 |
5.0 ± 0.13 |
| SA | 0.015% | 65.40 ± 0.59 |
6.28 ± 0.28 |
15.55 ± 0.48 |
9.65 ± 0.28 |
78.10 ± 0.49 |
12.7 ± 0.32 |
7.2 ± 0.19 |
4.7 ± 0.10 |
| SA | 0.020% | 60.25 ± 1.05 |
5.15 ± 0.15 |
17.95 ± 0.81 |
10.60 ± 0.14 | 73.10 ± 0.41 |
12.1 ± 0.29 |
6.9 ± 0.17 |
4.4 ± 0.08 |
| SA | 0.025% | 65.90 ± 0.61 |
6.20 ± 0.30 |
15.35 ± 0.49 |
9.60 ± 0.30 |
77.80 ± 0.52 |
12.4 ± 0.31 |
7.1 ± 0.18 |
4.6 ± 0.09 |
| Gamma rays | 10 kR | 87.50 ± 0.41 |
9.62 ± 0.21 |
8.75 ± 0.31 |
3.70 ± 0.33 |
89.10 ± 0.38 |
14.7 ± 0.37 |
8.9 ± 0.24 |
5.3 ± 0.14 |
| Gamma rays | 15 kR | 82.20 ± 0.62 |
8.18 ± 0.36 |
10.85 ± 0.22 |
4.05 ± 0.11 |
83.95 ± 0.70 |
14.1 ± 0.34 |
8.2 ± 0.21 |
5.1 ± 0.12 |
| Gamma rays | 20 kR | 79.10 ± 1.03 |
7.28 ± 0.55 |
12.75 ± 0.30 |
4.85 ± 0.35 |
78.00 ± 0.98 |
13.0 ± 0.32 |
7.6 ± 0.18 |
4.8 ± 0.10 |
| Gamma rays | 25 kR | 77.40 ± 1.15 |
6.05 ± 0.42 |
13.92 ± 0.47 |
6.45 ± 0.40 |
76.00 ± 0.69 |
12.3 ± 0.29 |
7.0 ± 0.17 |
4.5 ± 0.09 |
Values are mean ± SE (n = 3). Germination and survival percentages were calculated based on total seeds sown. Leaf morphological mutations include alterations in leaf shape, size and margin. Chlorophyll chimeras represent chlorophyll-deficient sectors observed in M₁ seedlings.
Table 2: Effect of EMS, sodium azide and gamma irradiation on M₁ germination, growth, mutation frequencies and yield-related traits in kidney bean variety ‘Waghya’
| Treatment | Dose | Seed germination (%) | Seedling height (cm) | Leaf morph. changes (%) | Chlorophyll chimeras (%) | Plant survival (%) | Pod length (cm) | Pods/plant | Seeds/pod |
| Control | — | 84.10 ± 0.82 | 12.08 ± 0.51 | — | — | 85.45 ± 0.88 | 16.4 ± 0.39 | 9.3 ± 0.28 | 5.3 ± 0.12 |
| EMS | 0.10% | 74.20 ± 0.61 | 8.12 ± 0.26 | 11.25 ± 0.43 | 2.75 ± 0.51 | 82.10 ± 0.44 | 15.0 ± 0.36 | 8.3 ± 0.24 | 5.0 ± 0.10 |
| EMS | 0.15% | 72.60 ± 1.18 | 7.05 ± 0.16 | 13.70 ± 0.31 | 4.28 ± 0.26 | 78.00 ± 0.40 | 14.2 ± 0.35 | 7.7 ± 0.22 | 4.8 ± 0.10 |
| EMS | 0.20% | 67.45 ± 0.65 | 6.00 ± 0.05 | 17.05 ± 0.27 | 5.32 ± 0.66 | 74.20 ± 0.33 | 13.1 ± 0.32 | 7.0 ± 0.21 | 4.6 ± 0.09 |
| EMS | 0.25% | 74.65 ± 0.56 | 8.18 ± 0.25 | 11.15 ± 0.40 | 2.60 ± 0.54 | 82.40 ± 0.41 | 14.6 ± 0.36 | 7.9 ± 0.23 | 5.0 ± 0.11 |
| SA | 0.010% | 69.45 ± 1.10 | 6.90 ± 0.04 | 11.85 ± 0.21 | 5.60 ± 0.38 | 78.20 ± 0.50 | 14.3 ± 0.35 | 8.1 ± 0.24 | 4.9 ± 0.10 |
| SA | 0.015% | 65.80 ± 0.85 | 6.00 ± 0.14 | 14.40 ± 0.30 | 6.95 ± 0.46 | 74.70 ± 0.55 | 13.5 ± 0.33 | 7.5 ± 0.22 | 4.7 ± 0.09 |
| SA | 0.020% | 60.35 ± 0.56 | 5.08 ± 0.11 | 16.60 ± 0.20 | 8.05 ± 0.52 | 70.10 ± 0.70 | 12.7 ± 0.31 | 6.9 ± 0.20 | 4.5 ± 0.09 |
| SA | 0.025% | 66.20 ± 0.86 | 6.05 ± 0.14 | 14.10 ± 0.29 | 6.78 ± 0.42 | 74.90 ± 0.57 | 13.1 ± 0.34 | 7.2 ± 0.21 | 4.6 ± 0.09 |
| Gamma rays | 10 kR | 79.50 ± 0.96 | 9.62 ± 0.25 | 10.25 ± 0.52 | 2.05 ± 0.10 | 82.35 ± 0.92 | 15.3 ± 0.37 | 8.6 ± 0.25 | 5.1 ± 0.11 |
| Gamma rays | 15 kR | 76.20 ± 1.40 | 8.19 ± 0.42 | 12.40 ± 0.57 | 3.40 ± 0.32 | 81.40 ± 0.60 | 14.5 ± 0.36 | 8.0 ± 0.23 | 4.9 ± 0.10 |
| Gamma rays | 20 kR | 71.20 ± 1.82 | 7.10 ± 0.63 | 14.60 ± 0.54 | 5.38 ± 0.55 | 75.80 ± 0.57 | 13.7 ± 0.34 | 7.3 ± 0.22 | 4.7 ± 0.10 |
| Gamma rays | 25 kR | 76.10 ± 1.42 | 8.20 ± 0.43 | 12.45 ± 0.58 | 3.40 ± 0.33 | 81.40 ± 0.63 | 14.0 ± 0.35 | 7.8 ± 0.23 | 4.8 ± 0.10 |
Values are mean ± SE (n = 3). Germination and survival percentages were calculated based on total seeds sown. Leaf morphological mutations include alterations in leaf shape, size and margin. Chlorophyll chimeras represent chlorophyll-deficient sectors observed in M₁ seedlings.
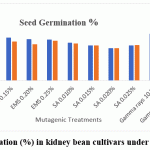 |
Figure 1: Seed germination (%) in kidney bean cultivars under mutagenic treatments. |
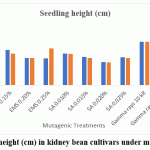 |
Figure 2: Seedling height (cm) in kidney bean cultivars under mutagenic treatments. |
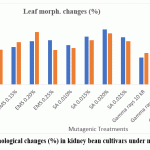 |
Figure 3: Leaf morphological changes (%) in kidney bean cultivars under mutagenic treatments. |
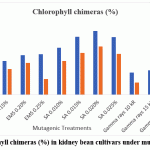 |
Figure 4: Chlorophyll chimeras (%) in kidney bean cultivars under mutagenic treatments. |
Discussion
The present study showed that gamma rays, EMS and sodium azide affected germination, seedling growth, mutation frequency and yield-related traits in two kidney bean cultivars. All mutagens caused dose-dependent reductions in germination and seedling vigour while increasing the frequency of leaf morphological mutations and chlorophyll chimeras, confirming the high sensitivity of common bean to induced mutagenesis.6,8,9
Sodium azide was the most damaging treatment, producing the strongest reductions in germination, seedling height and survival. This greater toxicity is consistent with the metabolic conversion of sodium azide to mutagenic intermediates such as azidoalanine, which can interfere with both nuclear DNA replication and mitochondrial function, leading to pronounced phytotoxicity and high frequencies of chlorophyll‑deficient sectors.12,14 EMS also exerted strong effects, particularly at 0.20%, where mutation frequency was high and seedling growth suppression was pronounced. As a monofunctional alkylating agent, EMS predominantly induces GC→AT base transitions via ethylation of guanine, which can disrupt genes controlling cell division, elongation and chlorophyll biosynthesis.8,13,20
Gamma rays were comparatively less damaging than the chemical mutagens at the doses used, although they still produced clear dose‑dependent reductions in growth and yield traits. Gamma irradiation primarily causes ionization and single‑ or double‑strand DNA breaks, resulting in chromosomal aberrations and altered cell division patterns. The relatively milder but still evident effects of gamma rays suggest that they may be useful for generating broader chromosomal variation with somewhat lower levels of physiological injury than the stronger chemical treatments, in agreement with other studies on legumes and beans.9,10,15
The two cultivars showed distinct responses to mutagenic stress. Varun exhibited higher chlorophyll mutation frequencies and more pronounced morphological variation, indicating greater mutability, whereas Waghya maintained better survival and more stable yield‑related traits across treatments. Such genotype‑dependent differences in mutagen sensitivity and mutability have been reported in several legume species,8-10 and reflect underlying variation in DNA repair capacity, genome organization and physiological tolerance. These findings highlight the importance of choosing mutagen doses that are tailored to the specific genetic background when designing mutation breeding programmes.
Yield-related traits were adversely affected by mutagenic treatments, with pod length and pod number being more strongly reduced than seeds per pod. This pattern suggests that early reproductive processes such as flower formation and pod development are more sensitive to mutagen-induced damage than the final number of seeds formed per pod.16-18 Nevertheless, some lower gamma rays doses & EMS preserved near-control levels of yield components while still inducing measurable mutation frequencies, indicating that these doses may offer a favourable balance between biological damage and useful variability.
Seed size and seed coat colour variations observed in the M₁ generation were irregular and non‑heritable, consistent with the expectation that M₁ primarily reflects physiological disturbance, chimeric sectors and temporary phenotypic changes. Stable seed coat colour mutants and other heritable morphological variants are more reliably detected and confirmed in M₂ and later generations, where segregation and fixation can be observed.9,19 Therefore, the present M₁ results should be regarded as preliminary indicators of mutagenic activity rather than a direct basis for selecting breeding lines.
In addition to the reductions observed in growth and yield-related traits, several distinct macro-mutants were identified in both kidney bean cultivars following mutagenic treatments. In Varun, mutants exhibiting dwarf, tall, luxuriant, robust, early flowering, large leaf, short pod, high-yielding, cream seed coat and blackish-red seed coat phenotypes were observed. In Waghya, dwarf, tall, small leaf, branched, broad pod/bold seed, high-yielding, early maturing, red seed coat, brown seed coat and shiny seed coat mutants were recorded. The occurrence of these diverse phenotypes indicates that gamma rays, EMS and sodium azide were effective in generating morphological variability in both cultivars. These mutants may serve as valuable genetic resources for future crop improvement; however, their stability and heritability should be confirmed through evaluation in the M₂ and subsequent generations before their potential use in breeding programmes is established.8,9
Overall, the findings demonstrate that induced mutagenesis can create valuable phenotypic variation in kidney bean, particularly when using EMS at 0.20% and sodium azide at 0.020–0.025%, which showed high mutation frequencies in both cultivars. However, the practical utility of this variation for crop improvement must be validated through careful screening and selection in M₂ and subsequent generations, as recommended in mutation breeding studies on legumes.8,9
Conclusion
Mutagenic treatments influenced germination, seedling growth, survival and yield related traits in kidney bean cultivars Varun and Waghya. Sodium azide produced the strongest inhibitory effects, while EMS at 0.20% and SA at 0.020–0.025% were the most effective in inducing leaf morphological mutations and chlorophyll chimeras. Varun exhibited higher mutability, particularly for chlorophyll mutations, whereas Waghya showed greater tolerance and stability in reproductive traits. The observed changes in seed coat colour and seed size in M₁ were transient and are best regarded as physiological responses rather than stable genetic mutations.
The present study demonstrates that induced mutagenesis is a useful strategy for generating variability in kidney bean, but the findings should be considered preliminary. Selection of stable, heritable mutants with desirable agronomic traits must be carried out in the M₂ and later generations before any breeding recommendations are made, in line with established mutation breeding practices in legumes.
Acknowledgement
The authors acknowledge the Head, Department of Botany, Ahmednagar College, Ahmednagar, for laboratory facilities and encouragement. We thank the Department of Biophysics, Government Institute of Science, Aurangabad, for gamma irradiation services, and the technical and field staff for their assistance.
Funding Sources
The author(s) received no financial support for the research, authorship, and/or publication of this article.
Conflict of Interest
The authors do not have any conflict of interest.
Data Availability Statement
This statement does not apply to this article.
Ethics Statement
This research did not involve human participants, animal subjects, or any material that requires ethical approval.
Informed Consent Statement
This study did not involve human participants, and therefore, informed consent was not required.
Clinical Trial Registration
This research does not involve any clinical trials.
Permission to reproduce material from other sources
Not Applicable
Author Contributions
- Dnyaneshwar Arjun Tuwar: Contributed to the conceptualization of the study, experimental design, execution of field and laboratory work, data collection, statistical analysis, interpretation of results, and preparation of the manuscript.
- Abhijit Arun Kulkarni: Contributed to the supervision of the research work, guidance in experimental planning, critical review and editing of the manuscript, and validation of results.
References
- Beebe S, Rao IM, Blair MW, Acosta‑Gallegos JA. Phenotyping common beans for adaptation to drought. Front Physiol. 2013;4:35. doi:10.3389/fphys.2013.00035
CrossRef - Costa GEA, Queiroz‑Monici KDS, Reis SMPM, Oliveira AC. Chemical composition, dietary fibre and resistant starch contents of raw and cooked bean (Phaseolus vulgaris L.) varieties. J Food Compos Anal. 2006;19(4):372‑379.
- Messina V. Nutritional and health benefits of dried beans. Am J Clin Nutr. 2014;100(Suppl 1):437S‑442S. doi:10.3945/ajcn.113.071472
CrossRef - Petry N, Boy E, Wirth JP, Hurrell R. The potential of the common bean (Phaseolus vulgaris) as a vehicle for iron biofortification. Nutrients. 2014;6(10):4357‑4384. doi:10.3390/nu6104357
- Broughton WJ, Hernández G, Blair MW, Beebe S, Gepts P, Vanderleyden J. Beans (Phaseolus spp.): model food legumes. Plant Soil. 2003;252(1):55‑128. doi:10.1023/A:1024146710611
CrossRef - Mudibu J, Nkongolo KK, Kizungu RV, Thine C. Genetic analysis and improvement of common bean (Phaseolus vulgaris L.) through induced mutagenesis. Int J Plant Breed Genet. 2012;6(4):214‑220.
- Varshney RK, Pandey MK, Bohra A, Singh VK, Thudi M, Saxena RK. Toward sequence‑based breeding in legumes. Biotechnol Adv. 2021;39:107461. doi:10.1016/j.biotechadv.2019.107461
CrossRef - Singh S, Datta SK. Induced mutagenesis in legumes: mechanisms and applications for crop improvement. Legume Res. 2019;42(3):345‑352. doi:10.18805/LR‑3962
- Wani MR. Mutation breeding in legumes: a review. J Appl Nat Sci. 2017;9(4):2146‑2154. doi:10.31018/jans.v9i4.1502
CrossRef - Laskar RA, Khan S. Genotypic response of legumes to EMS and irradiation. Mutation Breeding Rev. 2017;33(2):87‑102.
- Chakraborti D, Roy A, Ghosh P. Induced mutagenesis in French bean using EMS. J Genet. 2020;99:45‑53.
- Khan S, Wani AA, Parihar S. Mutagenic impact of sodium azide and EMS on germination traits in legumes. Int J Bot Stud. 2019;4(3):98‑105.
- Bharath R, Prasad BR, Hemalatha S. EMS‑induced variability in mung bean. Legume Res. 2023;46(2):215‑223.
- Pandey A, Gautam S, Singh B. Chlorophyll mutants as mutagenic indicators. J Radiat Res Appl Sci. 2018;11(2):163‑171.
- Verma A, Gupta R, Singh M. Gamma irradiation effects on survival and mutation frequency in beans. Radiat Bot. 2021;189:104‑113.
- Kumar R, Patil V. Chemical mutagen effects on reproductive traits in pulses. J Appl Biol Biotechnol. 2022;10(1):45‑51.
- Nabi G, Alam M, Yousuf S. Reproductive disruptions under mutagen stress in beans. Physiol Mol Biol Plants. 2023;29(1):167‑178.
- Shah T, Sher H, Khan MA. Ovule and seed formation stability in legumes under mutagen exposure. Euphytica. 2020;216(7):1‑12.
- Rehman F, Hussain W, Zafar Y. Seed coat color mutation patterns in legumes. J Crop Improv. 2019;33(4):503‑519.
- Sharma P, Singh C, Rana S. Oxidative stress responses of legumes to EMS. Plant Physiol Rep. 2021;26(3):423‑435.
Abbreviations
EMS – Ethyl Methanesulfonate
SA – Sodium azide