
Introduction
Agriculture has played a pivotal role in human civilization and food security. Plant breeding approaches have introduced improved crop varieties meeting the global food demand. However, the changing climate patterns have led to approximately a 20% reduction in the production of even the hybrid wheat varieties with the effects of drought being the most severe.1 To meet the needs of growing population, intensified agricultural practices led to a nutritionally depleted, less fertile and an unhealthy soil for the microflora to grow. The soil microflora is crucial in maintaining ecosystem dynamics and the role of rhizosphere microbiome in plant growth and yield cannot be over-looked. The rhizosphere harbouring several genes contribute to various traits and features exhibited by plant. Therefore, rhizoengengineering has emerged as a sustainable alternative to achieve food security maintaining soil health. The plant growth-promoting rhizobacteria (PGPR) act as miners, recyclers and refiners, providing nutrients and minerals to the plants.2 PGPR application has been effective in enhancing micronutrient (Mn, Fe, Co, Ni, Cu and Zn) as well as macronutrient (N, P, K, S) uptake in various plants. Utilization of PGPR in biofortification is not only encouraging in managing the challenges of malnutrition in populations but parallelly improves soil fertility, biodiversity and resilience of an ecosystem. The deficiencies of minerals and vitamins have affected a large population of the world leading to malnutrition. The diets of major countries are mostly the staple food crops and cereals often deficient in micronutrient. Soil and seedling inoculations is expanding in agribusiness as a helpful technique in manipulating microbial activity in the rhizosphere thereby improving plant growth and nutrient acquisition as depicted in Figure1. Seed biopriming helps in establishment of PGPR in the rhizosphere thereby enhancing the nutrient (Fe, P, N, Zn, and Cu) biofortification in cereal grains, through nutrient mobilization, exopolysaccharides and siderophores production. Acinetobacter, Bacillus, Brevibacterium, Enterobacter and Pseudomonas, are equally potent in biofortification as well as disease biocontrol in plants.3 The increasing solubility of micronutrients by these microorganisms are promising as they get incorporated in the edible plant parts like seeds ensuring an ideal food security. This further ensures an individual gets access to adequate food of right dietary mix. The key crops for micronutrient biofortification include wheat, rice, maize, sorghum and millets.
Deficiency of micronutrient makes people more vulnerable to physical and mental disorders, a feature more common in lower income countries having poor dietary diversity. On an average 149.2 million children under the age of five are estimated to be stunted and deaths in children under five due to malnutrition stands about 45 percent.4 Most of micronutrients consumed by human come from plant or animal products and on an average most of the Indian soils are deficient in zinc, iron, copper, manganese and boron. Micronutrient deficiencies in humans can be reduced through biofortification of crops. Increasing nutrients through rhizobacterial engineering is sustainable as well as acceptable by the consumers. Oats have become a preferred cereal with time due to its high fiber, proteins and low cholesterol content showing an annual production of 22 million tonnes.5 Experiments were therefore undertaken to check the effects of bacterial seed biopriming on oat grain nutrient fortification at different water levels.
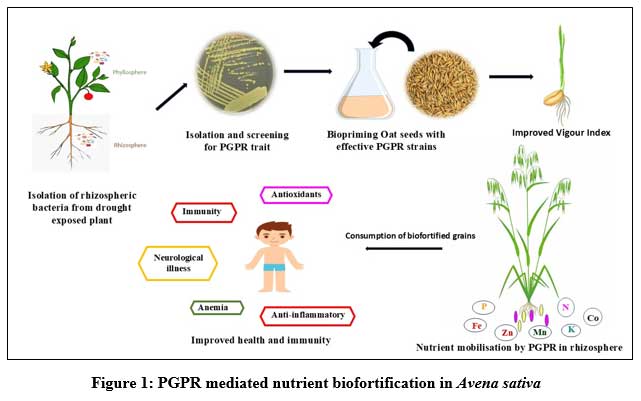 |
Figure 1: PGPR mediated nutrient biofortification in Avena sativa |
Materials and Methods
The study was conducted in the campus of BHU, Varanasi, India with a tropical monsoon climate. The experimental site was located at 25.2820 N latitude and 82.950 E longitudes, at the mean elevation of 76.9m above the sea level having silty loam soil with low permeability.
Seed biopriming and experimental design
Selected potential rhizospheric bacterial isolates, viz Enterobacter cloacae BHUSR1 (Accession No: MG913373), Serratia marcescens BHUSR2 (Accession No: MG913374) and Klebsiella aerogenes BHUSR3 (Accession No: MG913375), earlier tested and reported8 for their plant growth-promoting characters were tested for their effects on nutrient mobilisation in oat grains. The bacterial isolates were cultured on yeast extract mannitol (YEM) medium and were revived at every two-week intervals to maintain viability. For the purpose of seed inoculation, an adhesive solution was prepared by boiling gum acacia and sugar in distilled water for approximately 15 minutes and was allowed to cool. Certified seeds of oat (Avena sativa L.) cv. Harita RO-19, procured from the National Seed Research Centre, Varanasi, were surface disinfected and rinsed thoroughly with sterile water. The sterilized seeds were then immersed in individual bacterial suspensions for 7–8 hours to ensure proper coating. After incubation, the seeds were air-dried overnight under aseptic conditions in a laminar airflow cabinet and subsequently sown under field conditions. The experiments were carried out in a randomized complete block design (RCBD) with three replicates per treatment. Plot size for each treatment was 1.5m×3m and bioprimed seeds were sown manually by hand drilling in rows with and an inter row spacing of 45cm. After 30 days of germination the experimental plots were subjected to water stress (WS) by maintaining irrigation at intervals of 7 and 10 days, whereas for no water stress (NS), the plots were irrigated every 3 days or as required. Control sets without bacterial treatment were maintained at each irrigation level. Fifteen days after germination, seedlings were thinned to maintain a plant-to-plant spacing of 30 cm. Weeding was performed as needed throughout the crop growth period. The crop was harvested after 150 days of sowing and grains from each treatment plot were collected for nutrient analysis.
Nitrogen estimation
The nitrogen content of the grain samples was estimated using the Kjeldahl method. For estimation, 0.5 g of finely powdered grain sample was placed in a digestion tube, and 10 mL of a di-acid mixture (H2SO4:HClO4 in a 9:1 ratio) was added. The mixture was left overnight for pre-digestion. Subsequently, 10 g of a digestion mixture consisting of potassium sulfate and catalyst (CuSO4 and selenium powder) was added. The samples were digested in a digestion chamber until a clear, colorless solution was obtained. After cooling, the digest was filtered through Whatman No. 42 filter paper and collected in a volumetric flask, and the volume was adjusted to 20 mL with distilled water.
For distillation, 10 mL of 4% boric acid solution containing bromocresol green and methyl red indicators was taken in a conical flask, and the delivery outlet of the distillation unit was immersed in this solution. An aliquot of 5 mL of the digested sample was transferred to the distillation tube of a Kjeltec semi-automatic nitrogen analyzer, followed by the addition of 10 mL NaOH. The mixture was distilled for 9 minutes. The liberated ammonia was trapped in the boric acid solution and subsequently titrated against 0.02N H2SO4. A reagent blank was processed similarly.
Phosphorous estimation
Phosphorus content in oat seed samples was determined following the method described by Jackson.6 Phosphorous content in grain was calculated using standard curve prepared for 0, 2, 4, 6 and 8 μg ml-1.
Available potassium
The potassium content was estimated by the method described by Jackson.6 The photometer analysis was based on the measurement of the intensity of characteristic line emission given by the element to be determined.
Available zinc, iron and manganese in Avena sativa grain
Available micronutrient content in grain was estimated by Lindsay and Norvell.7 Grain samples were extracted for micronutrient analysis using DTPA and Mehlich-3 extractants. For extraction, 10 g of grain was shaken with 20 mL of DTPA solution (0.005 M DTPA, 0.01 M CaCl2, and 0.1 M triethanolamine, adjusted to pH 7.3) for 2 h, while 2 g of grain was extracted with 20 mL of Mehlich-3 solution (0.2 M acetic acid, 0.25 M ammonium nitrate, 0.015 M ammonium fluoride, 0.013 M nitric acid, and 0.001 M DTPA) for 5 min. The suspensions were filtered through Whatman No. 2 filter paper, and the filtrates were analyzed for Fe, Mn, and Zn using inductively coupled plasma (ICP) spectrometry at wavelengths of 238.204, 257.610, and 206.200 nm, respectively. The ICP was operated in radial plasma mode and calibration was performed using standard solutions of 0, 1, and 3 mg L⁻¹ prepared separately for each element.
Statistical analysis
Experiments were conducted in triplicate. The data were subjected to one-way analysis of variance (ANOVA), and mean comparisons were performed using Duncan’s Multiple Range Test (DMRT) at a significance level of p ≤ 0.05 using SPSS software (version 16.0). Correlation were performed by using Origin pro software 2024b.
Results
Rhizospheric microbiomes are crucial in plant growth and yield. Their activity in this region determines the availability of nutrients and therefore the quality and quantity of grains. In the present study the effect of seed biopriming with water stress tolerant PGPR’s E. cloacae BHUSR1, S. marcescens BHUSR2 and K. aerogenes BHUSR3 was checked on Avena sativa grain quality and quantity. Drought stress drastically reduced the growth and development of oat plants as reflected by reduced height, less biomass and wilted leaves. However, seed biopriming provided enough protection from water stress as all the selected PGPR’s were well adapted, being isolated from drought exposed areas and were good EPS producers as well.8 Seed biopriming significantly enhanced yield (Table 1). The highest grain yield (882 g plot⁻¹) was recorded in plots inoculated with K. aerogenes, representing a 54.29% increase over the uninoculated control under non-stress conditions. Improvements in yield was even observed after exposure to water stress, bringing 99% (7days) and 93% (10days) higher yields after biopriming. Among all the treated bacteria K. aerogenes performed best and enhanced yield at both water levels.
Table 1: Avena sativa grain yield after different bacterial treatments exposed to different water levels.
| S. No. | Treatments | Yield (g) | ||
| NS | WS7 | WS10 | ||
| 1. | Control | 571.66d | 390b | 296.66a |
| 2. | Enterobacter cloacae BHUSR1 | 818.33f | 752e | 494.33cd |
| 3. | Serratia marcescens BHUSR2 | 807.33f | 636d | 435c |
| 4. | Klebsiella aerogenes BHUSR3 | 882g | 776.66e | 572d |
(NS-No stress), (WS7-water stress of 7 days) and (WS10-water stress of 10 days). Value depicted is means of three replicates with different letters indicating significant differences among treatments according to DMRT at P ≤ 0.05
Water stress of 10 days brought significant decrease in the grain nutrient levels which was found to be compensated by the bacterial biopriming. The different bacteria were able to mobilize different nutrients from the soil with no common trend being observed. Under 7 and 10-days of water stress conditions, seed biopriming enhanced nitrogen accumulation in grains with the highest increase being observed in E. cloacae treated plants (28.30% and 30.27%), followed by K. aerogenes (23% and 25.48%) and S. marcescens (19.87% and 19.57%) when compared with their respective controls (Fig. 2A).The order of performance of strains was same even when the fields were normally irrigated with percent improvement in nitrogen content observed in E. cloacae (18.78%), K. aerogenes(17.74%) and S. marcescens (16.94%) with respect to their control. The nitrogen content in grains got improved and was recorded upto 7100 μg/kg after biopriming with E. cloacae when irrigated normally. In case of P-uptake, K. aerogenes performed best improving it by 51% followed by S. marcescens (36.73%) and E. cloacae (33.40%) when irrigated normally. After holding irrigation for 7 and 10 days the highest phosphorus content in the seeds was recorded in K. aerogenes (47.54 and 29.17%) followed by S. marcescens (34.74 and 24.89%) and E. cloacae (24.55 and 25.73%) when compared to their control (Fig. 2B). Significant effect of drought was observed on the K-content of seeds with the 10 days water stress exposure dropping down the K-content in seeds upto 45.26% when compared to the irrigated control plant seeds. Upto 34-39% enhancement in K-content was observed in most of the PGPR treated plants when normally irrigated with the bacterial effects being less specific. After 7 days of water stress the increment of seed K-content observed was S. marcescens (44%), K. aerogenes (34%) and E. cloacae (38%). Similarly, after 10 days stress exposure the percent K enhancement ranged from 48-39% with performance order being the same (Fig. 2C).
Zn is a vital micronutrient, enhancing seed weight. E. cloacae, K. aerogenes and S. marcescens being good zinc solubilizers were helpful in increasing the grain Zn content and 1000 grain weight. Biopriming followed by normal irrigation brought magnificent enhancement in Zn content of oat seed recorded maximum in K. aerogenes (50.49%) followed by S.marcescens (43.59%) and E. cloacae (37.36%). After 7 and 10 days of water stress conditions, highest Zn content (Fig. 2D) in seed was recorded in K. aerogenes (72.94 and 95.36%) followed by S. marcescens (60.33 and 84%) and E. cloacae (52.20 and 72.42%). Augmentation in grain Zn-content was many folds higher in water stressed fields compared to irrigated field grains for all the PGPR.
Iron deficiency can limit plant growth reducing crop yields, affecting seed quality as this element is essentially required in photosynthesis and its imbalance leads to chlorosis. Further iron forms the prosthetic group of many enzymes activating metabolic processes. The selected PGPR were good in siderophore production and mobilized iron well. Under normal irrigation highest iron content in seeds was recorded in wheat treated with K. aerogenes (44.19%) followed by E. cloacae (35.65%) and S. marcescens (34.11%) treated plants as compare to control. Introducing water stress of 7 and 10 days augmentation the Fe content in seeds of E. cloacae (35.11 and 49.33%) and K. aerogenes (25.37 and 48.54%) inoculated plants (Fig. 2E). S. marcescens inoculants did not have much effect on iron mobilization after water stress exposure.
Bacterial inoculations bringing significant improvement in manganese content were more effective under normal irrigation compared to water stressed conditions. Drought exposure brought a sharp decline in Mn-content. However, this could be overcome after seed biopriming by any of the selected PGPR strains. Mn-content after 7-days of stress was higher than the well-irrigated control plants. The bacterial performance were not very satisfactory on further increasing the stress duration to 10 days, although the difference was significant when compared to the respective non-primed drought exposed controls. Under no stress high content of soluble Mn in seed was recorded in K. aerogenes (41.64%), S. marcescens (29.28%) and E. cloacae (18.46%) while lowest was recorded in control plants (Fig. 2F). After 7 and 10 days of water stress the percentage Mn content declined in oat seed with the order being K. aerogenes (48.75 and 33.70%), S. marcescens (41.33 and 17.04%) and E. cloacae (36.62 and 20.51%).
 |
Figure 2: Effect of bacterial seed biopriming on macro and micronutrients contents in grains of Avena sativa under no stress (NS), water stress of 7 days (WS7) and 10 days (WS10). |
(A) Nitrogen content (B) Phosphorous content (C) Potassium content (D) Zinc content (E) Iron content (F) Manganese content. Results are mean of three replicates with different letters indicating significant differences among treatments according to DMRT at P≤0.05.
Correlation
 |
Figure 3: Pearson’s correlation matrix between Yield (Yl) and grain nutrient (N, P, K, Fe, Zn and Mn) at different water levels. (A) No stress (B) Water stress for 7 days (C) Water stress for 10 days. |
Under irrigated conditions yield was positively correlated to most of the nutrient content in grain (Fig. 3). Under 7 days water stress, the yield was much influenced by other nutrients compared to nitrogen, while under 10-days it was Fe and Mn that affected the yield more. The Zn content in grains was positively correlated to the Mn uptake. Similarly, the P-content in grains was positively correlated to the K, Zn and Mn contents irrespective of the water levels. Over all the effect of water stress on nutrient mobilization was clearly evident in the study. No significant correlation was found among the nutrient uptake when exposed to 10 days of drought.
Discussion
Management of soil lands affected by water scarcity is essential to meet the ever-enhancing food demands under changing climatic conditions. Plants’ ability to absorb nutrients like N, P, K, are negatively impacted by soil drought stress. Correa9 have revealed that drought stress lowers soil nutrient uptake by plants and decreases the concentration of N, K, and P in plant tissue. The current work aimed to check the effect of seed biopriming on nutrient uptake by plants highlights the complex interrelation of plants, PGPR and soil in influencing both macro and micronutrients content in Avena sativa grains grown under drought stress. Efficient strains have the ability to mobilise the fixed soil nutrients and acquisition, supporting plant development, also maintaining the soil health. Results of the experiments showed that inoculation with K. aerogenes, E. cloaca and S. marcescens significantly increased the K, N, P, Zn, Fe and Mn content in the oat seeds as compared to the controls that were uninoculated. It is well established that PGPR not only stimulate root growth but also enhance nutrient availability by solubilizing essential nutrients and activating ion transport systems within plant roots.10 This results in improved uptake of both macro and micronutrients as also observed in the present study. Nutrient uptake from the rhizosphere is the first step in this process, followed by translocation to shoots and eventual accumulation in grains.
Even when grown under water stressed conditions, the oat seeds inoculated with E. cloacae, S. marcescens and K. aerogenes exhibited higher nutrient content and were almost comparable to the plants grown under irrigated conditions. PGPR exhibits vital activities in soil including the fixation of nitrogen and solubilization of potassium and phosphate aiding in development and nutrient upliftment in plants.11 Under stress conditions such as drought, nutrient uptake is often limited due to reduced mass flow of water-soluble nutrients and decreased activity of key enzymes like nitrate reductase.12 However, PGPR inoculation helps mitigate these effects by modulating enzyme secretion and phytohormone levels thereby improving root system architecture. Increased root surface area and root tip density, enhances water and nutrient uptake under stress.13 Studies have shown that rhizobacteria such as Azospirillum can modulate endogenous IAA levels in wheat, improving plant water status and growth under stress conditions14,15 which correlates with the results observed in oat plants after bacterial biopriming.
Nitrogen (N) is an important and the primary component of amino acids building proteins and chlorophyll. Numerous non-protein substances, including co-enzymes, pigments that harvest light, secondary metabolites, and polyamines, have nitrogen as their components. Both energy production and metabolite operations in cells are impacted by nitrogen availability. Endophytes have a significant impact on the defense and plant survival mechanisms. Maximum nitrogen content in grains was improved by E. cloacae followed by K. aerogenes and S. marcescens in both drought and irrigated conditions as compared to their respective control. Bacillus is known for its ability to supply nitrogen in Cabbage plants.16 Co-inoculation with Pseudomonas putida NUU8 and Bradyrhizobium japonicum USDA strain significantly boosted the N, P, and K under drought conditions compared to the control.17 PGPR colonize the rhizosphere of the plant, stick to the root surface, and maintain moisture content in the rhizosphere through exopolysaccharide production.18 They also create stable aggregates that support plants’ uptake of nutrients.19 Seed biopriming was also helpful in increasing the phosphorus content of oat grains as compared the non-primed ones. Phosphorus is an essential element of nucleic acids, cellular membranes, and enzymes. It is needed for numerous cellular functions like photosynthesis, glucose metabolism, redox-homeostasis, and signaling. Phosphorus regulates water content, overcoming the adverse effects of salts on wheat plants, activating over 60 different plant enzymes. In terms of nutrition, abundant literature reports mycorrhizaas the most prevalent and significant symbiotic interaction between soil-dwelling fungi and roots helpful in phosphorus uptake.20 However, the PGPR too significantly contribute in phosphorus uptake through secretion of organic acid and phosphatases that mobilize phosphorous.21 In the present study too increased phosphorus content was observed in Klebsiella aerogenes plants followed by S. marcescens and E. cloacae when irrigated normally. Enhancement in uptake of nitrogen and phosphorous in wheat by the application of S. marcescens and Enterobacter sp. has also been reported earlier.22,23
Potassium is a macronutrient required for plant growth and development. It acts as a cofactor for more than 40 enzymes and a cation in establishing cell turgor and maintaining cell electroneutrality.24 Wheat inoculated plants showed an increase in the potassium content at both irrigation levels. Under normal conditions maximum increase in K-content was observed in K. aerogenes plants and under irrigation stress of either 7 or 10-days, high K increment was recorded in S. marcescens plants. Rani25 working with a native strain of Bacillus pseudomycoides, capable of solubilizing potassium increased its acquisition in tea seedlings grown in northeastern Indian soil treated with mica debris. Compared to control, the inoculation of Burkholderia sp. in plants of Mikania micrantha resulted in increased P and K uptake. Rahnella aquatilis, Pantoea agglomerans and Pseudomonas orientalis are three potassium-solubilizing bacteria whose effects on mobilization and uptake of K, P, and N by Mikania micrantha as studied by Sun.26 It was observed that the absorption of P was positively impacted by a sufficient supply of N, and P-starvation had a deleterious impact on N uptake and assimilation indicating a close association in their uptake.27 Synergistic effects of N and P uptake have been shown by Zhang28 to provide a substantially higher yield under varied environments. Enough availability of the necessary mineral nutrients such as K, N and P must be guaranteed for various physiological, metabolic, and biochemical processes to take place as intended in order for a crop plant to properly enter its reproductive phase. Nitrate (N) serves as a signal molecule to influence processes in addition to being a nutrient needed for the synthesis of starch and amino acids. Priming seed with PGPR caused a substantial increase in iron content of oat grains at both water levels. Maximum iron uptake was recorded in K. aerogenes and E. cloacae inoculated plants indicating a relatively healthier plant state as compared to the non-inoculated ones. Proven that the chosen PGPRs are well-known8 for their efficiency in mobilizing nutrients (P, Fe, and Zn), it is quite probable that they improved nutrient uptake even under stressful circumstances, increasing crop output and grain nutrient quality. Microorganisms have developed specialized processes for assimilating iron, such as the synthesis of low molecular weight compounds known as siderophores, which are helpful in delivering iron in plants. PGPRs ability to produce siderophores is crucial for biocontrol and the mobilization of iron through mineral chelation. The bioavailability of iron in soil is essential for a range of metabolic processes in plants, including oxygen transport, chloroplast development, chlorophyll biosynthesis, and mitochondrial functioning.29 Deficiency of iron in plants leads to chlorosis reducing crop production. Iron deficiency in grains is also a main cause of anemia in humans.
PGPR not only enhance the uptake of macronutrients30 but also influence micronutrient mobilization thereby improving plant growth and productivity as reported earlier in Avena sativa inoculated with Serratia and Erwinia.31 In addition, these bacteria influence overall plant physiology by improving root activity and nutrient transport efficiency. Zinc (Zn) is an essential element, and its deficiency can have adverse effects on human health. Over thirty percent of the global population suffers from zinc deficiency, affecting growth, compromised immune systems, skin issues, appetite loss, and hair loss, male hypogonadism, and delayed sexual maturation.32 Low persistency and high reactivity make the plant available Zn fraction very low in soil. Zn deficiency in plants leads to retarded root and shoot growth, affecting water and nutrient transport, chlorosis, decreased leaf size and improper pollen development leading to poor grain formation thereby affecting yield drastically.33 Novel methodologies for augmentation of zinc in crops include microbial strains inoculation in crops. Among microorganism both bacteria and fungi are reported to improve plant zinc availability in the rhizosphere and enhance zinc in cereals.34 Zn solubilizers include the bacterial genera Azospirillum, Azotobacter, Gluconacetobacter, Bacillus, and Pseudomonas.35 Kumar36 reported that arbuscular mycorrhizae and Trichoderma are the two major groups of fungi possessing Zn-solubilizing activities in soil. Our results confirm that seed biopriming with E. cloacae, S. marcescens and K. aerogenes are helpful in improving Zn-content in oat grains. These microbes essentially produce organic acids that help in sequestering Zn2+ that are subsequently absorbed by the crop plants. Zinc is involved in the activation of numerous enzymes required in metabolism of carbohydrates, proteins and lipids. Similarly, Mn content was augmented in grains undergone K. aerogenes treatment. Exposure to water stress declined the Mn content in oat grains as compared to irrigated ones. Mn is important in photolysis of water in chloroplasts, regulation of enzymatic activities and protection of membranes against oxidative damage as it is a cofactor that activates over 35 different types of enzymes in plants including superoxide dismutase and the Mn-protein of photosystem II.37 Most of these enzymes catalyze hydrolytic reactions, decarboxylation, and oxidation-reduction processes. Numerous cereals, including wheat, oats, sorghum, barley, rice, and legumes, have been reported to be equally sensitive to the levels of soil Mn either high or low.37 Manganese is also beneficial in biotic stress as it impacts plant resistance against root and foliar diseases, including blast rice, tomato wilt, common scab of potato, powdery mildew and downy mildew. Mn is crucial for photosynthesis, synthesis of secondary metabolites like phenol and lignin, as well as a few other physiological processes in plants that lead to the development of disease resistance.38 Lack of manganese inhibits photosynthetic activity, reduces the amount of lignin and other structural carbohydrates, and eventually hinders the growth of plants.39 According to Raghuwanshi37 Pseudomonas fluorescens and Bacillus subtilis when inoculated into crops under Mn deficiency increases Mn uptake, which enhances the formation of lignin and strengthens the crops’ defenses against certain phytopathogens. Some more studies reveal that by increasing the availability of Mn, Bacillus cereus enhanced the seed size, biomass, nutrient value of seed and overall yield.40 It is also found that certain actinomycetes, Streptomyces members, and several bacteria from the Pseudomonas and Bacillus genera increase the amount of Mn in crops41 as also observed in the Oat’s grain yield. Microbial inoculation of maize with potential PGPR strains i.e. Bacillus aryabhattai ZM31, Bacillus subtilis ZM63, Bacillus aryabhattai S10 and Paenibacillus polymyxa ZM27 improved plant development by uplifting the mineral nutrients in shoots and grains of maize.
Biopriming Avena sativa seeds with strains of PGPRs viz Klebsiella aerogenes Enterobacter cloacae and Serratia marcescens that were water stress tolerant significantly increased the growth of oat by improving its physiological and metabolic processes as reported earlier8. Improvement in crop yield after microbial inoculations under water stress is mainly associated with the enhanced root activity. All the three bacterial strains selected were good IAA producer8 which resulted in increased root length and hence the plants were able to explore a greater soil volume, thereby improving their performance even under suboptimal growth conditions, as also observed in other studies done on wheat treated with botanicals42 and oat inoculated by Klebsiella spp.37 PGPR stimulate plant growth and development through multiple mechanisms, primarily by modulating phytohormone levels and enhancing nutrient availability in the rhizosphere. One of the key mechanisms involves the production of phytohormones such as auxins (indole-3-acetic acid, IAA), which regulate root system architecture, including cell division, elongation, and vascular differentiation. This expanded root system improves the plant’s ability to absorb water and nutrients, ultimately promoting plant growth and development.
Uptake of macro as well as micro-nutrients was found to be positively correlated with each other affecting yield. Factors affecting nutrient uptake are mostly dependent on soil pH, nutrient availability, root architecture and microbial activity in the rhizosphere. The selected microbes capable of regulating the soil pH and hormones were effective in managing nutrient uptake promoting yield. A positive relationship between nutrient content (N, P, Fe) and plant biomass was also recorded in earlier studies1. In our results, PGPR inoculation led to a significant increase in grain yield, along with enhanced concentrations of essential nutrients such as N, P, K, Fe, Zn, and Mn in grains. This improvement can be attributed to increased nutrient availability in the soil due to PGPR activity, which enhanced nutrient uptake by roots. Subsequently, these nutrients were efficiently translocated from roots to aerial parts and finally accumulated in the grains. Thus, the observed enrichment of nutrients in grains indicates improved nutrient uptake-translocation efficiency mediated by PGPR. Partitioning of nutrients especially nitrogen and phosphorus responsible for growth and development at the early stages have been supportive as reported in maize1 and wheat.43 Further, the biofortification of Fe and Zn observed during the investigation accredits to the secretion of siderophores by these bacterial strains. Earlier studies too done on Avena sativa have reported increased nutrient uptake through mineral solubilization and siderophore production by Serratia and Erwinia31 biopriming. About more than three million people world-wide are under micronutrient deficiency and malnutrition. Enhancing the bioavailable nutrients in staple crops mediated by PGPR can be an affordable and easy option among the strategies targeting biofortification of micronutrients. The selected PGPR’s being good mineral solubilizers enhanced nutrient uptake even under stressed conditions there by improving the grain quality. Overall, the observed improvement in oat plant growth can be accredited to the bacterial mediated nutrient mobilization and uptake.
Conclusion
Micronutrient deficiency has significant adverse effects on grain quality and yield also exposing them to biotic stress. The selected rhizospheric bacterial strains capable of mobilizing soil nutrient were helpful in improving the nutrient uptake and their partitioning in grains biofortifying micronutrients. Improvement in nutrients content and yield of oat was observed after deploying selected drought tolerant rhizospheric bacterial strains at different water levels. Among all the bacterial strains Klebsiella aerogenes BHUSR3 was the most effective in fortifying nutrients in oat grains. Thus, biofortification through seed biopriming can be an easy, cost-effective strategy even under varied water levels expanding the cultivation to marginal lands. This strategy holds strong potential for applications in sustainable agriculture, especially in areas with limited-resources and high risk of drought. Further field trials in different agro-climatic conditions may strengthen the finding. The challenges of synergism of introduced microbes with native microbes and limited shelf life of biofertilizers are concerns to be addressed.
Acknowledgment
The authors are grateful to Banaras Hindu University for providing the necessary facilities for the laboratory and field work.
Funding Sources
Sandeep Gupta would like to acknowledge the financial support as fellowship, received from UGC, Government of India New Delhi. Authors are thankful to the Banaras Hindu University, IoE grant 6031, for providing the necessary facilities and support.
Conflict of Interest
The authors do not have any conflict of interest.
Data Availability Statement
All data generated or analyzed during this study are included in this published article. All data to support the conclusions have been either provided or are otherwise publicly available.
Ethics Statement
This research did not involve human participants, animal subjects, or any material that requires ethical approval.
Informed Consent Statement
This study did not involve human participants, and therefore, informed consent was not required.
Clinical Trial Registration
This research does not involve any clinical trials.
Permission to reproduce material from other sources
Not Applicable
Author Contributions
Richa Raghuwanshi, Sandeep Gupta: Conceptualization.
Sandeep Gupta, Seema Devi, Jay Kishor Prasad, Riddha Dey, Surya Prakash Dube: Formal analysis and investigation.
Seema Devi, Poonam Singh: Writing.
Richa Raghuwanshi, Riddha Dey, Surya Prakash Dube: Review and editing.
Richa Raghuwanshi: Funding acquisition, Resources, Supervision
References
- Ahmad M., Hussain A., Dar A., Luqman M., Ditta A., Iqbal Z., Ahmad H.T., Nazli F., Soufan W., Almutairi K., Sabagh A. E. Combating iron and zinc malnutrition through mineral biofortification in maize through plant growth promoting Bacillus and Paenibacillus species. Front. Plant Sci, 2023; 13,1094551.https://doi.org/10.3389/fpls.2022.1094551
CrossRef - Argentel-Martínez L., Peñuelas-Rubio O., Herrera-Sepúlveda A., González-Aguilera J., Sudheer S., Salim L. M., Lal S., Pradeep C. K., Ortiz K., Sansinenea E., Hathurusinghe S. H. K., Shin J. H., Babalola O. O., Azizoglu U. Biotechnological advances in plant growth-promoting rhizobacteria for sustainable agriculture. World J. Microbiol. Biotechnol, 2025; 41(1), 21. https://doi.org/10.1007/s11274-024-04231-4
CrossRef - Dube S. P., Dey R., Devi S., Raghuwanshi R. Seed Quality, Chlorophyll and Carotene Content in Brassica juncea L. Leaves at Two Growth Stages in Response to Rhizospheric Bacteria. Curr. Agric. Res. J, 2024; 12(2). http://dx.doi.org/10.12944/CARJ.12.2.25
CrossRef - World Health Organization. Monitoring health for the SDGs, sustainable development goals. World Health Water Air Soil Pollut Organization. Retrived from https://www.amazon. com/World-Health-Statistics-2016-Sustainable/dp/9241565268.
- Jalal A., Oliveira C. E. D. S., Gato I. M. B., Moreira V. D. A., de Lima B. H., Bastos A. D. C., Teixeira Filho M. C. M. Interaction of mineral nutrients and plant growth-promoting microbes for biofortification of different cropping systems. J. Plant Growth Regul, 2025; 44(12), 6785-6801. https://doi.org/10.1007/ s00344-024-11380-1
CrossRef - Jackson M. Soil chemical analysis. Prentice Hall. Inc., Englewood Cliffs, NJ, 1958; 498, 183-204. https://doi.org/10.1002/jpln.19590850311
CrossRef - Kolokotsa A., Koukoulakis P. H., Kyritsis S. S., Kalavrouziotis I. K. The Impact of Heavy Metal Interactions with pH, Organic Matter and Clays on Soil Toxicity, Under Wastewater and Biosolid Reuse. Water Air Soil Pollut, 2025; 236(7), 412.https://doi.org/10.1007/s11270-025-08069-w
CrossRef - Gupta S. K., Dey R., Devi S., Raghuwanshi R. Implications and adaptive responses in Avena sativa towards rhizospheric bacteria under drought stress. The Microbe, 2024; 100159.https://doi.org/10.1016/j.microb.2024.100159
CrossRef - Correa S. S., Pacheco R. S., Viana G. C., Vidal M. S., Xavier G. R., de Araújo J. L. S. Ability of nitrogen-fixing bacteria to alleviate drought stress in cowpea varies depending on the origin of the inoculated strain. Plant Soil, 2024; 498(1), 391-408. https://doi.org/10.1007/s11104-023-06443-3
CrossRef - Carreiras J., Cruz-Silva A., Fonseca B., Carvalho R. C., Cunha J. P., Proença Pereira, J. Improving grapevine heat stress resilience with marine plant Growth-Promoting rhizobacteria consortia. Microorganisms, 2023; 11 (4). https://doi.org/10.3390/microorganisms11040856
CrossRef - Zahoor I., Farzana M. H. Ch., Shafeeq M., Afzal A. B., Haider S. M. S., Arshad E., Zafar M., Ahmad S. Enhancing Soil Nutrient Efficiency for Sustainable Agriculture. Sch J Agric Vet Sci, 2025; 7, 232-241. https://doi.org/10.36347/sjavs.2025.v12i07.001
CrossRef - Li S., Zhou L., Addo-Danso S. D., Ding G., Sun M., Wu S., Lin, S. Nitrogen supply enhances the physiological resistance of Chinese fir plantlets under polyethylene glycol (PEG)-induced drought stress. Scientific Reports, 2020; 10(1), 7509. https://doi.org/10.1038/s41598-020-64161-7
CrossRef - Ortiz-Castro R., Campos-García J., López-Bucio J. Pseudomonas putida and Pseudomonas fluorescens influence Arabidopsis root system architecture through an auxin response mediated by bioactive cyclodipeptides. Plant Growth Regul, 2020; 39(1), 254-265. https://doi.org/10.1007/s00344-019-09979-w
CrossRef - Singh T. B., Sahai V., Ali A., Prasad M., Yadav A., Shrivastav P., Goyal D., Dantu P. K. Screening and evaluation of PGPR strains having multiple PGP traits from hilly terrain. J Appl Biol Biotechnol, 2020; 8(4), 38-44. https://doi.org/10.7324/JABB.2020.80406
CrossRef - Karimi N., Goltapeh E. M., Amini J., Mehnaz S., Zarea M. J. (2021). Effect of Azospirillum zeae and seed priming with zinc, manganese and auxin on growth and yield parameters of wheat, under dryland farming. Agric Res, 2021;10(1), 44-55. https://doi.org/10.1007/s40003-020-00480-5
CrossRef - Liu Z., Yu B., Xu Y., Yang S., Cang J., Peng Y., Tan J., Liu L., Li W., Liu X., Wei M. Bacillus-functionalized sewage sludge biochar boosts cabbage growth through improved nitrogen assimilation. Biochar, 2026; 8(1), 42. https://doi.org/10.1007/s42773-025-00561-0
CrossRef - Hassen A. I., Muema E. K., Diale M. O., Mpai T., Bopape F. L. Non-rhizobial endophytes (NREs) of the nodule microbiome have synergistic roles in beneficial tripartite plant–microbe interactions. Microorganisms, 2025; 13(3), 518. https://doi.org/10.3390/microorganisms13030518
CrossRef - Raghuwanshi R. Prospects of cropping with polysaccharides producing microbes under drought stress. Res. J. Agric. Sci, 2024; 55(1), 2-10.https://doi.org/10.5152/AUAF.2024.23114
CrossRef - Wang M., Xu Z. PGPR-mediated enhancement of soil nutrients, rhizosphere microbial ecology, and plant growth: a review. npj Biofilms and Microbiomes, 2026. https://doi.org/10.1038/s41522-026-00966-0
CrossRef - Chaudhary A., Poudyal S., Kaundal A. Role of arbuscular mycorrhizal fungi in maintaining sustainable agroecosystems. J. Appl. Microbiol, 2025; 5(1), 6. https://doi.org/10.3390/applmicrobiol5010006
CrossRef - Lei Y., Kuai Y., Guo M., Zhang H., Yuan Y., Hong H. Phosphate-solubilizing microorganisms for soil health and ecosystem sustainability: A forty-year scientometric analysis (1984–2024). Front. Microbiol, 2025; 16, 1546852. http://dx.doi.org/10.3389/fmicb.2025.1546852
CrossRef - Gupta S., Dube S. P., Raghuwanshi R. Rhizoengineering influences on soil dynamics and microbial communities in post-harvest oat fields. Soil Adv, 2025; 3, 100032. https://doi.org/10.1016/j.soilad. 2024.100032
CrossRef - Marchese S. I., Canchero J. O., Puig N., Curá J. A., Miralles D. J. Yield increases through Azospirillum spp. and Enterobacter spp. inoculations in wheat. Cereal Res. Commun, 2025; 53(2), 1105-1117. https://doi.org/10.1007/s42976-024-00571-x
CrossRef - Wegner L. H., Pottosin I., Dreyer I., Shabala S. Potassium homeostasis and signalling: from the whole plant to the subcellular level. Quant. Plant Bio, 2025; 6, e13. https://doi.org/10.1017/qpb.2025.10
CrossRef - Rani K., Biswas D. R., Basak B. B., Bhattacharyya R., Biswas S., Das T. K., Bandyopadhyay K. K., Kaushik R., Das A., Thakur J. K., Agarwal B. K. Exploring waste mica as an alternative potassium source using a novel potassium solubilizing bacterium and rice residue in K deficient Alfisol. Plant Soil, 2025; 509(1), 611-630. https://doi.org/10.1007/s11104-024-06879-1
CrossRef - Sun F., Ou Q., Wang N., xuan Guo Z., Ou Y., Li N., Peng C. Isolation and identification of potassium-solubilizing bacteria from Mikania micrantha rhizospheric soil and their effect on M. micrantha plants. Glob. Ecol. Conserv, 2020; 23, e01141. https://doi.org/10.1016/j.gecco.2020.e01141
CrossRef - Pandey R., Sharma S., Mishra A., Sakhare A. S., Meena S. K., Vengavasi K. Dual-nutrient stress tolerance in wheat is regulated by nitrogen and phosphorus uptake, assimilation, reutilization, and differential expression of candidate genes. Plant Soil, 2025; 508(1), 117-142. https://doi.org/10.1007/s11104-024-06789-2
CrossRef - Zhang Y. J., Si Y. L., Ju S. B., Jiang W. Y., Zhao M. Z., Liu J., Tigabu M., Ma X. Q., Li M. Synergistic effects of nitrogen deposition and phosphorus distribution patterns on root morphological and chemical traits, and nutrient efficiency in Chinese fir. J. Plant Ecol, 2025; 18(4), rtaf062. https://doi.org/10.1093/jpe/rtaf062
CrossRef - Gu S., Wang N., Zheng Y., Wang T., Shen Q., Zhang F., Kümmerli R., Wei Z., Zuo Y. Integrating microbial siderophores into concepts of plant iron nutrition. Nat. Plants, 2025; 1-11. https://doi.org/10.1038/s41477-025-02171-x
CrossRef - Behera B., Das T. K., Raj R., Ghosh S., Raza M. B., Sen, S. Microbial consortia for sustaining productivity of non-legume crops: prospects and challenges. Agric Res, 2021; 10(1), 1-14. https://doi.org/10.1007/s40003-020-00482-3
CrossRef - Devi R., Alsaffar M. F., AL-Taey D. K., Kumar S., Negi R., Sharma B., Kaur T., Rustagi S., Kour D., Yadav A.N., Ahluwalia A. S. Synergistic effect of minerals solubilizing and siderophores producing bacteria as different microbial consortium for growth and nutrient uptake of oats (Avena sativa). Vegetos, 2024; 37(5), 1863-1875. https://doi.org/10.1007/s42535-024-00922-3
CrossRef - Gadapati S. The Public Health Significance of Human Zinc Deficiency in the Developing World. In Strategies to Alleviate Human Zinc Deficiency (pp. 49-64). IGI Global Scientific Publishing. 2025. https://doi.org/10.4018/979-8-3693-5355-4.ch003
CrossRef - Majumdar A., Saraf S. K., Sahu C., Pathak P., Baghel M. Zinc Deficiency: Impact on Growth, Immunity, and the Role of Plant‐Based Nutritional Strategies. Chem. Biodivers, 2025; 22(10), e00325. https://doi.org/10.1002/cbdv.202500325
CrossRef - Suganya A., Saravanan A. Effect of Zn fertilizer in combinations with arbuscular mycorrhizal fungi and zinc solubilising bacteria on micronutrients transformations and uptake in maize grown on calcareous soils. Maize J, 2025; 4000, 54.
- Rani N., Chauhan A., Kaur S., Solanki M. K., Tripathi M., Jain D., Singh S., Upadhyay S. K., Kaur G. Molecular mechanistic of Zn-solubilizing bacteria for agronomic eminence: recent updates and futuristic development. J. Plant Growth Regul, 2025; 44(4), 1337-1351. https://doi.org/10.1007/s00344-023-11111-y
CrossRef - Kumar M., Narwal E., Rakshit S., Bishi S. K., Bhadana V. P., Kumar M., Lal S. K., Monika., Jha U. C., Roy S., Layek J. Zinc homeostasis and biofortification in crop plants: towards efficient zinc utilization. J. Plant Growth Regul, 2025; 44(10), 5926-5945. https://doi.org/10.1007/s00344-025-11805-5
CrossRef - Raghuwanshi R., Gupta S. K., Prasad J. K., Dey R., Devi S., Dube S. P., Tripathi Y. N. Uptake of Micro-Nutrients by Plants through Microbial Channels: A Review. Curr. Agric. Res. J, 2025; 13(2). http://dx.doi.org/10.12944/CARJ.13.2.1
CrossRef - Jeevanraj R., Sivakumar R., Boominathan P., Kavitha P. S., Sendhilvel V. The essential role of macronutrients and micronutrients in improving crop resilience to biotic stress. J. Plant Physiol, 2025; 72(4), 126. https://doi.org/10.1134/S1021443724608838
CrossRef - Li Y., Fu Y., Gan Z., Wei Q., Yang M., Yao F., Zhou G. Manganese deficiency exacerbates boron deficiency-induced corky split vein in citrus by disrupting photosynthetic physiology and enhancing lignin metabolism. Horticulturae, 2025; 11(10), 1172. https://doi.org/10.3390/horticulturae11101172
CrossRef - Bhardwaj I., Bhardwaj N., Kumar V., Kumari S., Dulta K., Aman J., Onyeaka H., Ghosh S. Synergistic Effects of Bacillus cereus Mn‐5 PGPR‐Derived Silver Oxide Nanoparticles on Tomato Plant Growth, Stress Resilience and Nutritional Enhancement. J. Sustain. Agric. Environ, 2025; 4(4), e70090. https://doi.org/10.1002/sae2.70090
CrossRef - Djebaili R., Farda B., Gialdini O., Vaccarelli I., Rezaee Danesh Y., Pellegrini M. Microbial Consortium of Streptomyces spp. from Mining Environments Enhances Phytoremediation Potential of Lemna minor L. Plants, 2025; 14(22), 3467. https://doi.org/10.3390/plants14223467
CrossRef - Devi S., Tripathi Y. N., Raghuwanshi R. Himalayan botanical mediated elicitation regulates wheat growth through secondary metabolites and hormonal signaling. Agric. Biotechnol, 2025; 103851. https://doi.org/10.1016/j.bcab.2025.103851
CrossRef - Prasad J. K., Dey R., Raghuwanshi R. Exopolysaccharide-Producing rhizospheric bacteria enhance yield via promoting wheat (Triticum aestivum) growth at early stages. Microbiology, 2022; 91(6), 757-769. https://doi.org/10.1134/S0026261721102622
CrossRef
Abbreviations List
IAA: Indole acetic acid
NS: No stress
PGPR: Plant growth-promoting rhizobacteria
WS: Water stress
YEM: Yeast extract mannitol